ಪತ್ರರಂಧ್ರ
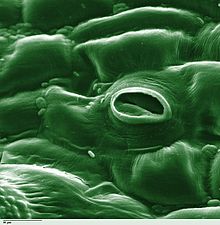


ಸಸ್ಯಶಾಸ್ತ್ರದಲ್ಲಿ, ಸ್ಟೋಮಾ ( ಗ್ರೀಕ್ನಿಂದ στόμα, "ಬಾಯಿ", ಬಹುವಚನ "ಸ್ಟೊಮಾಟಾ"), ಇದನ್ನು ಸ್ಟೊಮೇಟ್ (ಬಹುವಚನ "ಸ್ಟೊಮೇಟ್ಸ್") ಎಂದೂ ಕರೆಯುತ್ತಾರೆ. ಇದು ಎಲೆಗಳು, ಕಾಂಡಗಳು ಮತ್ತು ಇತರ ಅಂಗಗಳ ಎಪಿಡರ್ಮಿಸ್ನಲ್ಲಿ ಕಂಡುಬರುವ ಒಂದು ರಂಧ್ರವಾಗಿದೆ. ಅದು ಅನಿಲ ವಿನಿಮಯ ದರವನ್ನು ನಿಯಂತ್ರಿಸುತ್ತದೆ. ರಂಧ್ರವು ಪತ್ರರಂಧ್ರದ ತೆರೆಯುವಿಕೆಯ ಗಾತ್ರವನ್ನು ನಿಯಂತ್ರಿಸುವ ಜವಾಬ್ದಾರಿಯನ್ನು ಹೊಂದಿರುವ ಗಾರ್ಡ್ ಕೋಶಗಳೆಂದು ಕರೆಯಲ್ಪಡುವ ಒಂದು ಜೋಡಿ ವಿಶೇಷ ಪ್ಯಾರೆಂಚೈಮಾ ಕೋಶಗಳ ಗಡಿಯನ್ನು ಹೊಂದಿವೆ.
ಸಾಮಾನ್ಯವಾಗಿ ಜೋಡಿಯಾಗಿರುವ ಕಾವಲು ಕೋಶಗಳು ಮತ್ತು ರಂಧ್ರವನ್ನು ಒಳಗೊಂಡಿರುವ ಸಂಪೂರ್ಣ ಪತ್ರರಂಧ್ರದ ಸಂಕೀರ್ಣವನ್ನ ದ್ಯುತಿರಂಧ್ರ ಎಂದು ಕರೆಯಲಾಗುತ್ತದೆ.[೧] ಉಸಿರಾಟದಲ್ಲಿ ಬಳಸಲಾಗುವ ಆಮ್ಲಜನಕವನ್ನು ಹೊಂದಿರುವ ಗಾಳಿ ಮತ್ತು ದ್ಯುತಿಸಂಶ್ಲೇಷಣೆಯಲ್ಲಿ ಬಳಸಲಾಗುವ ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್, ಅನಿಲ ಪ್ರಸರಣದಿಂದ ಪತ್ರರಂಧ್ರದ ಮೂಲಕ ಹಾದುಹೋಗುತ್ತದೆ. ನೀರಿನ ಆವಿ ಸ್ಟೊಮಾಟಾದ ಮೂಲಕ ವಾತಾವರಣಕ್ಕೆ ಉತ್ಕರ್ಷಣ ಎಂಬ ಪ್ರಕ್ರಿಯೆಯಲ್ಲಿ ಹರಡುತ್ತದೆ.
ಲಿವರ್ವರ್ಟ್ಗಳನ್ನು ಹೊರತುಪಡಿಸಿ ಎಲ್ಲಾ ಭೂ ಸಸ್ಯ ಗುಂಪುಗಳ ಸ್ಪೊರೊಫೈಟ್ ಪೀಳಿಗೆಯಲ್ಲಿ ಸ್ಟೊಮಾಟಾ ಇರುತ್ತದೆ. ನಾಳೀಯ ಸಸ್ಯಗಳಲ್ಲಿ ಪತ್ರರಂಧ್ರದ ಸಂಖ್ಯೆ, ಗಾತ್ರ ಮತ್ತು ವಿತರಣೆಯು ವ್ಯಾಪಕವಾಗಿ ಬದಲಾಗುತ್ತದೆ. ಡೈಕೋಟಿಲ್ಡಾನ್ಗಳು ಸಾಮಾನ್ಯವಾಗಿ ಮೇಲಿನ ಮೇಲ್ಮೈಗಿಂತ ಎಲೆಗಳ ಕೆಳಗಿನ ಮೇಲ್ಮೈಯಲ್ಲಿ ಹೆಚ್ಚು ಪತ್ರರಂಧ್ರವನ್ನು ಹೊಂದಿರುತ್ತವೆ. ಈರುಳ್ಳಿ, ಓಟ್ ಮತ್ತು ಮೆಕ್ಕೆ ಜೋಳದಂತಹ ಮೊನೊಕೋಟಿಲ್ಡಾನ್ಗಳು ಎರಡೂ ಎಲೆಗಳ ಮೇಲ್ಮೈಗಳಲ್ಲಿ ಒಂದೇ ಸಂಖ್ಯೆಯ ಪತ್ರರಂಧ್ರವನ್ನು ಹೊಂದಿರಬಹುದು.[೨] ಟೆಂಪ್ಲೇಟು:Rpತೇಲುವ ಎಲೆಗಳನ್ನು ಹೊಂದಿರುವ ಸಸ್ಯಗಳಲ್ಲಿ, ಮೇಲ್ಭಾಗದ ಎಪಿಡರ್ಮಿಸ್ನಲ್ಲಿ ಮಾತ್ರ ಪತ್ರರಂಧ್ರ ಕಂಡುಬರಬಹುದು ಮತ್ತು ಮುಳುಗಿರುವ ಎಲೆಗಳು ಪತ್ರರಂಧ್ರವನ್ನು ಹೊಂದಿರುವುದಿಲ್ಲ. ಹೆಚ್ಚಿನ ಮರಗಳು ಎಲೆಯ ಕೆಳಗಿನ ಮೇಲ್ಮೈಯಲ್ಲಿ ಮಾತ್ರ ಪತ್ರರಂಧ್ರವನ್ನು ಹೊಂದಿರುತ್ತವೆ.[೩] ಎಲೆಗಳ ಮೇಲಿನ ಮತ್ತು ಕೆಳಗಿನ ಎರಡೂ ಮೇಲ್ಮೈಗಳಲ್ಲಿ ಪತ್ರರಂಧ್ರವನ್ನು ಹೊಂದಿರುವ ಎಲೆಗಳನ್ನು ಆಂಫಿಸ್ಟೊಮ್ಯಾಟಸ್ ಎಲೆಗಳು ಎಂದು ಕರೆಯಲಾಗುತ್ತದೆ; ಕೆಳಗಿನ ಮೇಲ್ಮೈಯಲ್ಲಿ ಮಾತ್ರ ಪತ್ರರಂಧ್ರವನ್ನು ಹೊಂದಿರುವ ಎಲೆಗಳು ಹೈಪೋಸ್ಟೊಮ್ಯಾಟಸ್ ಆಗಿರುತ್ತವೆ ಮತ್ತು ಮೇಲಿನ ಮೇಲ್ಮೈಯಲ್ಲಿ ಮಾತ್ರ ಪತ್ರರಂಧ್ರ ಹೊಂದಿರುವ ಎಲೆಗಳು ಎಪಿಸ್ಟೊಮ್ಯಾಟಸ್ ಅಥವಾ ಹೈಪರ್ಸ್ಟೊಮ್ಯಾಟಸ್ ಆಗಿರುತ್ತವೆ.[೩] ಪತ್ರರಂಧ್ರದ ಗಾತ್ರವು ಜಾತಿಗಳಾದ್ಯಂತ ಬದಲಾಗುತ್ತದೆ. ೧೦ ರಿಂದ ೮೦ µm ವರೆಗೆ ಉದ್ದ ಮತ್ತು ಅಗಲವು ೫೦ µm ರವರೆಗೆ ಅಂತ್ಯದಿಂದ ಅಂತ್ಯದವರೆಗೆ [೪]
ಕಾರ್ಯ

ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ ಲಾಭ ಮತ್ತು ನೀರಿನ ನಷ್ಟ
ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್, ದ್ಯುತಿಸಂಶ್ಲೇಷಣೆಯಲ್ಲಿ ಪ್ರಮುಖ ಪ್ರತಿಕ್ರಿಯಾಕಾರಿ. ಇದು ಸುಮಾರು ೪೦೦ ಪಿಪಿವಎಮ್ ಸಾಂದ್ರತೆಯಲ್ಲಿ ವಾತಾವರಣದಲ್ಲಿದೆ. ಹೆಚ್ಚಿನ ಸಸ್ಯಗಳಿಗೆ ಹಗಲಿನ ವೇಳೆಯಲ್ಲಿ ಪತ್ರರಂಧ್ರ ತೆರೆದಿರಬೇಕು. ಎಲೆಗಳಲ್ಲಿನ ಗಾಳಿಯ ಸ್ಥಳಗಳು ನೀರಿನ ಆವಿಯಿಂದ ಸ್ಯಾಚುರೇಟೆಡ್ ಆಗಿರುತ್ತವೆ, ಇದು ಟ್ರಾನ್ಸ್ಪಿರೇಶನ್ ಎಂದು ಕರೆಯಲ್ಪಡುವ ಪ್ರಕ್ರಿಯೆಯಲ್ಲಿ ಪತ್ರರಂಧ್ರದ ಮೂಲಕ ಎಲೆಯಿಂದ ನಿರ್ಗಮಿಸುತ್ತದೆ. ಆದ್ದರಿಂದ, ಸಸ್ಯಗಳು ಏಕಕಾಲದಲ್ಲಿ ನೀರಿನ ಆವಿಯನ್ನು ಕಳೆದುಕೊಳ್ಳದೆ ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ ಅನ್ನು ಪಡೆಯಲು ಸಾಧ್ಯವಿಲ್ಲ.[೫]
ಪರ್ಯಾಯ ವಿಧಾನಗಳು
ಸಾಮಾನ್ಯವಾಗಿ, ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ ಅನ್ನು ೧,೫-ಬಿಸ್ಫಾಸ್ಫೇಟ್ (RuBP) ಗೆ ರೈಬುಲೋಸ್ ೧,೫-ಬಿಸ್ಫಾಸ್ಫೇಟ್ (RuBP) ಗೆ ರುಬಿಸ್ಕೋ ಕಿಣ್ವದಿಂದ ಮೆಸೊಫಿಲ್ ಕೋಶಗಳಲ್ಲಿ ನೇರವಾಗಿ ಎಲೆಯೊಳಗಿನ ಗಾಳಿಯ ಸ್ಥಳಗಳಿಗೆ ಒಡ್ಡಲಾಗುತ್ತದೆ. ಇದು ಎರಡು ಕಾರಣಗಳಿಗಾಗಿ ಟ್ರಾನ್ಸ್ಪಿರೇಷನ್ ಸಮಸ್ಯೆಯನ್ನು ಉಲ್ಬಣಗೊಳಿಸುತ್ತದೆ: ಮೊದಲನೆಯದಾಗಿ, ರುಬಿಸ್ಕೋ ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ಗೆ ತುಲನಾತ್ಮಕವಾಗಿ ಕಡಿಮೆ ಸಂಬಂಧವನ್ನು ಹೊಂದಿದೆ, ಮತ್ತು ಎರಡನೆಯದಾಗಿ, ಇದು RuBP ಗೆ ಆಮ್ಲಜನಕವನ್ನು ಸರಿಪಡಿಸುತ್ತದೆ, ಫೋಟೊರೆಸ್ಪಿರೇಷನ್ ಎಂಬ ಪ್ರಕ್ರಿಯೆಯಲ್ಲಿ ಶಕ್ತಿ ಮತ್ತು ಇಂಗಾಲವನ್ನು ವ್ಯರ್ಥ ಮಾಡುತ್ತದೆ. ಈ ಎರಡೂ ಕಾರಣಗಳಿಗಾಗಿ, ರುಬಿಸ್ಕೋ ಗೆ ಹೆಚ್ಚಿನ ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ ಸಾಂದ್ರತೆಯ ಅಗತ್ಯವಿದೆ, ಅಂದರೆ ವಿಶಾಲವಾದ ದ್ಯುತಿರಂಧ್ರಗಳು ಮತ್ತು ಪರಿಣಾಮವಾಗಿ, ಹೆಚ್ಚಿನ ನೀರಿನ ನಷ್ಟ.
ಕಿರಿದಾದ ದ್ಯುತಿರಂಧ್ರಗಳನ್ನು ಹೆಚ್ಚಿನ ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ ಸಂಬಂಧದೊಂದಿಗೆ ಮಧ್ಯವರ್ತಿ ಅಣುವಿನ ಜೊತೆಯಲ್ಲಿ ಬಳಸಬಹುದು. ಫಾಸ್ಫೋನೊಲ್ಪೈರುವೇಟ್ ಕಾರ್ಬಾಕ್ಸಿಲೇಸ್ (PEPcase) ನಿಂದ ಕಾರ್ಬನ್ ಸ್ಥಿರೀಕರಣದ ಉತ್ಪನ್ನಗಳನ್ನು ಹಿಂಪಡೆಯುವುದು ಶಕ್ತಿ-ತೀವ್ರ ಪ್ರಕ್ರಿಯೆಯಾಗಿದೆ. ಪರಿಣಾಮವಾಗಿ, PEPCase ಪರ್ಯಾಯವು ನೀರು ಸೀಮಿತವಾಗಿದ್ದರೂ ಬೆಳಕು ಸಮೃದ್ಧವಾಗಿರುವಲ್ಲಿ ಮಾತ್ರ ಉತ್ತಮವಾಗಿರುತ್ತದೆ, ಅಥವಾ ಹೆಚ್ಚಿನ ತಾಪಮಾನವು ಕಾರ್ಬನ್ ಡೈಆಕ್ಸೈಡ್ಗೆ ಹೋಲಿಸಿದರೆ ಆಮ್ಲಜನಕದ ಕರಗುವಿಕೆಯನ್ನು ಹೆಚ್ಚಿಸುತ್ತದೆ, ರುಬಿಸ್ಕೋ ಆಮ್ಲಜನಕೀಕರಣದ ಸಮಸ್ಯೆಯನ್ನು ಹೆಚ್ಚಿಸುತ್ತದೆ.
ಸಿಎಎಮ್ ಸಸ್ಯಗಳು

"CAM" ಸಸ್ಯಗಳೆಂದು ಕರೆಯಲ್ಪಡುವ ಬಹುಮಟ್ಟಿಗೆ ಮರುಭೂಮಿ ಸಸ್ಯಗಳ ಗುಂಪು (ಕ್ರಾಸ್ಸುಲೇಸಿ ಕುಟುಂಬದ ನಂತರ ಸಿಎಎಮ್ ಪ್ರಕ್ರಿಯೆಯು ಮೊದಲು ಕಂಡುಹಿಡಿದ ಜಾತಿಗಳನ್ನು ಒಳಗೊಂಡಿರುವ ಕ್ರಾಸ್ಸುಲೇಸಿ ಆಸಿಡ್ ಮೆಟಾಬಾಲಿಸಮ್ ) ರಾತ್ರಿಯಲ್ಲಿ ತಮ್ಮ ಪತ್ರರಂಧ್ರವನ್ನು ತೆರೆಯುತ್ತದೆ (ನೀರು ಎಲೆಗಳಿಂದ ನಿಧಾನವಾಗಿ ಆವಿಯಾದಾಗ) ಕಾರ್ಬನ್ ಡೈಆಕ್ಸೈಡ್ ಅನ್ನು ಸರಿಪಡಿಸಲು ಮತ್ತು ಉತ್ಪನ್ನಗಳನ್ನು ದೊಡ್ಡ ನಿರ್ವಾತಗಳಲ್ಲಿ ಸಂಗ್ರಹಿಸಲು PEPಕಾರ್ಬಾಕ್ಸಿಲೇಸ್ ಅನ್ನು ಬಳಸಿ. ಮರುದಿನ, ಅವರು ತಮ್ಮ ಸ್ಟೊಮಾಟಾವನ್ನು ಮುಚ್ಚುತ್ತಾರೆ ಮತ್ತು ಹಿಂದಿನ ರಾತ್ರಿ ಸ್ಥಿರವಾದ ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ ಅನ್ನು ರುಬಿಸ್ಕೊ ಉಪಸ್ಥಿತಿಗೆ ಬಿಡುಗಡೆ ಮಾಡುತ್ತಾರೆ. ಇದು ರುಬಿಸ್ಕೊ ಅನ್ನು ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ನೊಂದಿಗೆ ಸ್ಯಾಚುರೇಟ್ ಮಾಡುತ್ತದೆ. ಇದು ಕನಿಷ್ಟ ದ್ಯುತಿಉತ್ಕರ್ಷವನ್ನು ಅನುಮತಿಸುತ್ತದೆ. ಆದಾಗ್ಯೂ, ಈ ವಿಧಾನವು ನಿರ್ವಾತಗಳಲ್ಲಿ ಸ್ಥಿರ ಇಂಗಾಲವನ್ನು ಸಂಗ್ರಹಿಸುವ ಸಾಮರ್ಥ್ಯದಿಂದ ತೀವ್ರವಾಗಿ ಸೀಮಿತವಾಗಿದೆ, ಆದ್ದರಿಂದ ನೀರು ತೀವ್ರವಾಗಿ ಸೀಮಿತವಾದಾಗ ಮಾತ್ರ ಇದು ಯೋಗ್ಯವಾಗಿರುತ್ತದೆ.
ಪತ್ರರಂಧ್ರ ತೆರೆಯುವುದು ಮತ್ತು ಮುಚ್ಚುವುದು

ಆದಾಗ್ಯೂ, ಹೆಚ್ಚಿನ ಸಸ್ಯಗಳು CAM ಅನ್ನು ಹೊಂದಿಲ್ಲ ಮತ್ತು ಆದ್ದರಿಂದ ಬೆಳಕಿನ ತೀವ್ರತೆ, ಆರ್ದ್ರತೆ ಮತ್ತು ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ ಸಾಂದ್ರತೆಯಂತಹ ಬದಲಾಗುತ್ತಿರುವ ಪರಿಸ್ಥಿತಿಗಳಿಗೆ ಪ್ರತಿಕ್ರಿಯೆಯಾಗಿ ಹಗಲಿನ ಸಮಯದಲ್ಲಿ ತಮ್ಮ ಪತ್ರರಂಧ್ರವನ್ನು ತೆರೆಯಬೇಕು ಮತ್ತು ಮುಚ್ಚಬೇಕು. ಪತ್ರರಂಧ್ರ ತೆರೆಯುವಿಕೆಗೆ ಪರಿಸ್ಥಿತಿಗಳು ಅನುಕೂಲಕರವಾದಾಗ (ಉದಾಹರಣೆಗೆ, ಹೆಚ್ಚಿನ ಬೆಳಕಿನ ತೀವ್ರತೆ ಮತ್ತು ಹೆಚ್ಚಿನ ಆರ್ದ್ರತೆ), ಪ್ರೋಟಾನ್ ಪಂಪ್ ಪ್ರೋಟಾನ್ಗಳನ್ನು (H + ) ಗಾರ್ಡ್ ಕೋಶಗಳಿಂದ ಓಡಿಸುತ್ತದೆ. ಇದರರ್ಥ ಜೀವಕೋಶಗಳ ವಿದ್ಯುತ್ ಸಾಮರ್ಥ್ಯವು ಹೆಚ್ಚು ಋಣಾತ್ಮಕವಾಗಿರುತ್ತದೆ. ಋಣಾತ್ಮಕ ವಿಭವವು ಪೊಟ್ಯಾಸಿಯಮ್ ವೋಲ್ಟೇಜ್-ಗೇಟೆಡ್ ಚಾನಲ್ಗಳನ್ನು ತೆರೆಯುತ್ತದೆ ಮತ್ತು ಆದ್ದರಿಂದ ಪೊಟ್ಯಾಸಿಯಮ್ ಅಯಾನುಗಳ (ಕೆ + ) ಹೀರಿಕೊಳ್ಳುವಿಕೆ ಸಂಭವಿಸುತ್ತದೆ. ಈ ಆಂತರಿಕ ನಕಾರಾತ್ಮಕ ವೋಲ್ಟೇಜ್ ಅನ್ನು ನಿರ್ವಹಿಸಲು ಪೊಟ್ಯಾಸಿಯಮ್ ಅಯಾನುಗಳ ಪ್ರವೇಶವು ನಿಲ್ಲುವುದಿಲ್ಲ, ಋಣಾತ್ಮಕ ಅಯಾನುಗಳು ಪೊಟ್ಯಾಸಿಯಮ್ನ ಒಳಹರಿವನ್ನು ಸಮತೋಲನಗೊಳಿಸುತ್ತವೆ. ಕೆಲವು ಸಂದರ್ಭಗಳಲ್ಲಿ, ಕ್ಲೋರೈಡ್ ಅಯಾನುಗಳು ಪ್ರವೇಶಿಸುತ್ತವೆ, ಆದರೆ ಇತರ ಸಸ್ಯಗಳಲ್ಲಿ ಸಾವಯವ ಅಯಾನು ಮಾಲೇಟ್ ಕಾವಲು ಕೋಶಗಳಲ್ಲಿ ಉತ್ಪತ್ತಿಯಾಗುತ್ತದೆ. ದ್ರಾವಕ ಸಾಂದ್ರತೆಯ ಈ ಹೆಚ್ಚಳವು ಜೀವಕೋಶದೊಳಗಿನ ನೀರಿನ ಸಾಮರ್ಥ್ಯವನ್ನು ಕಡಿಮೆ ಮಾಡುತ್ತದೆ, ಇದು ಆಸ್ಮೋಸಿಸ್ ಮೂಲಕ ಜೀವಕೋಶದೊಳಗೆ ನೀರಿನ ಪ್ರಸರಣಕ್ಕೆ ಕಾರಣವಾಗುತ್ತದೆ. ಇದು ಜೀವಕೋಶದ ಪರಿಮಾಣ ಮತ್ತು ಟರ್ಗರ್ ಒತ್ತಡವನ್ನು ಹೆಚ್ಚಿಸುತ್ತದೆ. ನಂತರ, ಸೆಲ್ಯುಲೋಸ್ ಮೈಕ್ರೊಫೈಬ್ರಿಲ್ಗಳ ಉಂಗುರಗಳಿಂದಾಗಿ ಕಾವಲು ಕೋಶಗಳ ಅಗಲವನ್ನು ಊತದಿಂದ ತಡೆಯುತ್ತದೆ ಮತ್ತು ಹೆಚ್ಚುವರಿ ಟರ್ಗರ್ ಒತ್ತಡವು ಗಾರ್ಡ್ ಕೋಶಗಳನ್ನು ವಿಸ್ತರಿಸಲು ಅನುವು ಮಾಡಿಕೊಡುತ್ತದೆ. ಅದರ ತುದಿಗಳನ್ನು ಸುತ್ತುವರಿದ ಎಪಿಡರ್ಮಲ್ ಕೋಶಗಳಿಂದ ದೃಢವಾಗಿ ಇರಿಸಲಾಗುತ್ತದೆ, ಎರಡು ಕಾವಲು ಕೋಶಗಳು ಉದ್ದವಾಗುತ್ತವೆ. ಪರಸ್ಪರ ದೂರವಾಗಿ ಬಾಗುವ ಮೂಲಕ, ತೆರೆದ ರಂಧ್ರವನ್ನು ರಚಿಸುವ ಮೂಲಕ ಅನಿಲವನ್ನು ಹರಡಬಹುದು.[೬]
ಬೇರುಗಳು ಮಣ್ಣಿನಲ್ಲಿ ನೀರಿನ ಕೊರತೆಯನ್ನು ಗ್ರಹಿಸಲು ಪ್ರಾರಂಭಿಸಿದಾಗ, ಅಬ್ಸಿಸಿಕ್ ಆಮ್ಲ (ಎಬಿಎ) ಬಿಡುಗಡೆಯಾಗುತ್ತದೆ.[೭] ಎಬಿಎ ಕಾವಲು ಕೋಶಗಳ ಪ್ಲಾಸ್ಮಾ ಮೆಂಬರೇನ್ ಮತ್ತು ಸೈಟೋಸೋಲ್ನಲ್ಲಿನ ಗ್ರಾಹಕ ಪ್ರೋಟೀನ್ಗಳಿಗೆ ಬಂಧಿಸುತ್ತದೆ, ಇದು ಮೊದಲು ಜೀವಕೋಶಗಳ ಸೈಟೋಸೋಲ್ನ ಪಿಎಚ್ ಅನ್ನು ಹೆಚ್ಚಿಸುತ್ತದೆ ಮತ್ತು ಕೋಶದ ಹೊರಗಿನ ಒಳಹರಿವು ಮತ್ತು ಎಂಡೋಪ್ಲಾಸ್ಮಿಕ್ ರೆಟಿಕ್ಯುಲಮ್ ಮತ್ತು ವ್ಯಾಕ್ಯೂಲ್ಗಳಂತಹ ಆಂತರಿಕ ಮಳಿಗೆಗಳಿಂದ Ca 2+ ಬಿಡುಗಡೆಯ ಕಾರಣದಿಂದಾಗಿ ಸೈಟೋಸಾಲ್ನಲ್ಲಿ Ca 2+ ಸಾಂದ್ರತೆಯನ್ನು ಹೆಚ್ಚಿಸುತ್ತದೆ..[೮] ಇದು ಕ್ಲೋರೈಡ್ (Cl - ) ಮತ್ತು ಸಾವಯವ ಅಯಾನುಗಳನ್ನು ಜೀವಕೋಶಗಳಿಂದ ನಿರ್ಗಮಿಸಲು ಕಾರಣವಾಗುತ್ತದೆ. ಎರಡನೆಯದಾಗಿ, ಇದು ಜೀವಕೋಶಗಳು ಮತ್ತಷ್ಟು K + ಅನ್ನು ಹೀರಿಕೊಳ್ಳುವುದನ್ನು ನಿಲ್ಲಿಸುತ್ತದೆ ಮತ್ತು ತರುವಾಯ, K + ನಷ್ಟವಾಗುತ್ತದೆ. ಈ ದ್ರಾವಕಗಳ ನಷ್ಟವು ನೀರಿನ ಸಾಮರ್ಥ್ಯದಲ್ಲಿ ಹೆಚ್ಚಳಕ್ಕೆ ಕಾರಣವಾಗುತ್ತದೆ, ಇದು ಆಸ್ಮೋಸಿಸ್ ಮೂಲಕ ಕೋಶದಿಂದ ಮತ್ತೆ ನೀರಿನ ಪ್ರಸರಣಕ್ಕೆ ಕಾರಣವಾಗುತ್ತದೆ. ಇದು ಕೋಶವನ್ನು ಪ್ಲಾಸ್ಮೋಲೈಸ್ ಮಾಡುತ್ತದೆ, ಇದು ಪತ್ರರಂಧ್ರ ಮುಚ್ಚಲು ಕಾರಣವಾಗುತ್ತದೆ.
ಎಪಿಡರ್ಮಲ್ ಕೋಶಗಳಿಂದ ಕಾವಲು ಕೋಶಗಳನ್ನು ಪಡೆಯಲಾಗುತ್ತದೆ.ಗಾರ್ಡ್ ಕೋಶಗಳು ಇತರ ಎಪಿಡರ್ಮಲ್ ಕೋಶಗಳಿಗಿಂತ ಹೆಚ್ಚು ಕ್ಲೋರೊಪ್ಲಾಸ್ಟ್ಗಳನ್ನು ಹೊಂದಿರುತ್ತವೆ, ಅವುಗಳ ಕಾರ್ಯವು ವಿವಾದಾಸ್ಪದವಾಗಿದೆ.[೯][೧೦]
ಅನಿಲ ವಿನಿಮಯದಿಂದ ಪತ್ರರಂಧ್ರ ನಡವಳಿಕೆಯನ್ನು ನಿರ್ಣಯಿಸುವುದು
ಎಲೆಯ ಅನಿಲ ವಿನಿಮಯವನ್ನು ಅಳೆಯುವ ಮೂಲಕ ಪತ್ರರಂಧ್ರ ಪ್ರತಿರೋಧದ ಮಟ್ಟವನ್ನು ನಿರ್ಧರಿಸಬಹುದು. ಟ್ರಾನ್ಸ್ಪಿರೇಶನ್ ದರವು ಪತ್ರರಂಧ್ರಗಳಿಂದ ಒದಗಿಸಲಾದ ಪ್ರಸರಣ ಪ್ರತಿರೋಧವನ್ನು ಅವಲಂಬಿಸಿರುತ್ತದೆ ಮತ್ತು ಎಲೆಯ ಆಂತರಿಕ ಗಾಳಿಯ ಸ್ಥಳಗಳು ಮತ್ತು ಹೊರಗಿನ ಗಾಳಿಯ ನಡುವಿನ ತೇವಾಂಶದ ಗ್ರೇಡಿಯಂಟ್ ಅನ್ನು ಅವಲಂಬಿಸಿರುತ್ತದೆ. ಪತ್ರರಂಧ್ರ ಪ್ರತಿರೋಧವನ್ನು (ಅಥವಾ ಅದರ ವಿಲೋಮ, ಪತ್ರರಂಧ್ರ ವಾಹಕತೆ ) ಆದ್ದರಿಂದ ಟ್ರಾನ್ಸ್ಪಿರೇಶನ್ ದರ ಮತ್ತು ಆರ್ದ್ರತೆಯ ಗ್ರೇಡಿಯಂಟ್ನಿಂದ ಲೆಕ್ಕ ಹಾಕಬಹುದು. ಇದು ಬೆಳಕಿನ ತೀವ್ರತೆ ಮತ್ತು ನೀರಿನ ಆವಿ, ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್ ಮತ್ತು ಓಝೋನ್ನಂತಹ ಅನಿಲಗಳ ಸಾಂದ್ರತೆಯಂತಹ ಪರಿಸರ ಪರಿಸ್ಥಿತಿಗಳಲ್ಲಿನ ಬದಲಾವಣೆಗಳಿಗೆ ಪತ್ರರಂಧ್ರ ಹೇಗೆ ಪ್ರತಿಕ್ರಿಯಿಸುತ್ತದೆ ಎಂಬುದನ್ನು ತನಿಖೆ ಮಾಡಲು ವಿಜ್ಞಾನಿಗಳಿಗೆ ಅನುವು ಮಾಡಿಕೊಡುತ್ತದೆ.[೧೧] ಬಾಷ್ಪೀಕರಣ ( E ) ಅನ್ನು ಹೀಗೆ ಲೆಕ್ಕ ಹಾಕಬಹುದು;[೧೨]
r
ಇಲ್ಲಿ e i ಮತ್ತು e a ಗಳು ಕ್ರಮವಾಗಿ ಎಲೆ ಮತ್ತು ಸುತ್ತುವರಿದ ಗಾಳಿಯಲ್ಲಿ ನೀರಿನ ಆಂಶಿಕ ಒತ್ತಡಗಳು, P ಎಂಬುದು ವಾತಾವರಣದ ಒತ್ತಡ, ಮತ್ತು r ಎಂಬುದು ಪತ್ರರಂಧ್ರದ ಪ್ರತಿರೋಧ. r ನ ವಿಲೋಮವು ನೀರಿನ ಆವಿಗೆ ವಾಹಕತೆಯಾಗಿದೆ ( g ), ಆದ್ದರಿಂದ ಸಮೀಕರಣವನ್ನು ಮರುಹೊಂದಿಸಬಹುದು;[೧೨]
ಮತ್ತು g ಗೆ ಪರಿಹರಿಸಲಾಗಿದೆ;[೧೨]
ದ್ಯುತಿಸಂಶ್ಲೇಷಕ CO 2 ಸಮೀಕರಣ ( A ) ನಿಂದ ಲೆಕ್ಕ ಹಾಕಬಹುದು
ಇಲ್ಲಿ C a ಮತ್ತು C i ಕ್ರಮವಾಗಿ CO 2 ರ ವಾಯುಮಂಡಲದ ಮತ್ತು ಉಪ-ಪತ್ರರಂಧ್ರದ ಭಾಗಶಃ ಒತ್ತಡಗಳು. ದ್ಯುತಿಸಂಶ್ಲೇಷಣೆ ವ್ಯವಸ್ಥೆಯನ್ನು ಬಳಸಿಕೊಂಡು ಎಲೆಯಿಂದ ಆವಿಯಾಗುವ ದರವನ್ನು ನಿರ್ಧರಿಸಬಹುದು. ಈ ವೈಜ್ಞಾನಿಕ ಉಪಕರಣಗಳು ಎಲೆಯಿಂದ ಹೊರಡುವ ನೀರಿನ ಆವಿಯ ಪ್ರಮಾಣವನ್ನು ಮತ್ತು ಸುತ್ತುವರಿದ ಗಾಳಿಯ ಆವಿಯ ಒತ್ತಡವನ್ನು ಅಳೆಯುತ್ತವೆ. ದ್ಯುತಿಸಂಶ್ಲೇಷಕ ವ್ಯವಸ್ಥೆಗಳು ನೀರಿನ ಬಳಕೆಯ ದಕ್ಷತೆ ( A/E ), g, ಅಂತರ್ಗತ ನೀರಿನ ಬಳಕೆಯ ದಕ್ಷತೆ ( A/g ) ಮತ್ತು C i ಅನ್ನು ಲೆಕ್ಕಾಚಾರ ಮಾಡಬಹುದು. ಈ ವೈಜ್ಞಾನಿಕ ಉಪಕರಣಗಳನ್ನು ಸಾಮಾನ್ಯವಾಗಿ ಸಸ್ಯ ಶರೀರಶಾಸ್ತ್ರಜ್ಞರು CO 2 ಹೀರಿಕೊಳ್ಳುವಿಕೆಯನ್ನು ಅಳೆಯಲು ಮತ್ತು ದ್ಯುತಿಸಂಶ್ಲೇಷಕ ದರವನ್ನು ಅಳೆಯಲು ಬಳಸುತ್ತಾರೆ.[೧೩][೧೪]
ವಿಕಾಸ
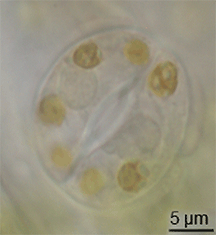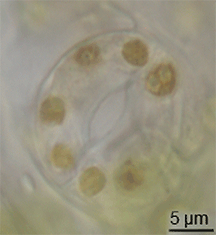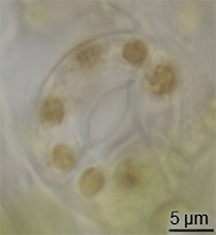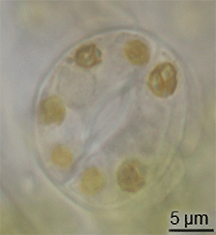
ಪಳೆಯುಳಿಕೆ ದಾಖಲೆಯಲ್ಲಿ ಸ್ಟೊಮಾಟಾದ ವಿಕಸನದ ಬಗ್ಗೆ ಕಡಿಮೆ ಪುರಾವೆಗಳಿವೆ, ಆದರೆ ಅವು ಸೈಲೂರಿಯನ್ ಅವಧಿಯ ಮಧ್ಯಭಾಗದಲ್ಲಿ ಭೂಮಿ ಸಸ್ಯಗಳಲ್ಲಿ ಕಾಣಿಸಿಕೊಂಡವು.[೧೫] ಸಸ್ಯಗಳ ಪಾಚಿ-ತರಹದ ಪೂರ್ವಜರಿಂದ ಪರಿಕಲ್ಪನೆಗಳನ್ನು ಮಾರ್ಪಡಿಸುವ ಮೂಲಕ ಅವು ವಿಕಸನಗೊಂಡಿರಬಹುದು.[೧೬] ಆದಾಗ್ಯೂ, ಪತ್ರರಂಧ್ರದ ವಿಕಸನವು ಮೇಣದ ಹೊರಪೊರೆ ವಿಕಸನಗೊಳ್ಳುವ ಸಮಯದಲ್ಲಿ ಸಂಭವಿಸಿರಬೇಕು - ಈ ಎರಡು ಗುಣಲಕ್ಷಣಗಳು ಒಟ್ಟಾಗಿ ಆರಂಭಿಕ ಭೂಮಿಯ ಸಸ್ಯಗಳಿಗೆ ಪ್ರಮುಖ ಪ್ರಯೋಜನವಾಗಿದೆ.ಟೆಂಪ್ಲೇಟು:Fact
ಅಭಿವೃದ್ಧಿ
ಮೂರು ಪ್ರಮುಖ ವಿಧದ ಎಪಿಡರ್ಮಲ್ ಕೋಶಗಳಿವೆ, ಇವೆಲ್ಲವೂ ಅಂತಿಮವಾಗಿ ಚಿಗುರಿನ ತುದಿಯ ಮೆರಿಸ್ಟೆಮ್ನ ಹೊರಗಿನ (L1) ಅಂಗಾಂಶ ಪದರದಿಂದ ಉತ್ಪತ್ತಿಯಾಗುತ್ತದೆ, ಇದನ್ನು ಪ್ರೊಟೊಡರ್ಮಲ್ ಕೋಶಗಳು ಎಂದು ಕರೆಯಲಾಗುತ್ತದೆ: ಟ್ರೈಕೋಮ್ಗಳು, ಪಾದಚಾರಿ ಕೋಶಗಳು ಮತ್ತು ಕಾವಲು ಕೋಶಗಳು, ಇವೆಲ್ಲವೂ ಯಾದೃಚ್ಛಿಕವಲ್ಲದ ಶೈಲಿಯಲ್ಲಿ ಜೋಡಿಸಲ್ಪಟ್ಟಿವೆ.
ಪ್ರೋಟೋಡರ್ಮಲ್ ಕೋಶಗಳಲ್ಲಿ ಅಸಮಪಾರ್ಶ್ವದ ಕೋಶ ವಿಭಜನೆಯು ಸಂಭವಿಸುತ್ತದೆ ಇದರ ಪರಿಣಾಮವಾಗಿ ಒಂದು ದೊಡ್ಡ ಕೋಶವು ಪಾದಚಾರಿ ಕೋಶವಾಗಿ ಪರಿಣಮಿಸುತ್ತದೆ ಮತ್ತು ಮೆರಿಸ್ಟೆಮೊಯ್ಡ್ ಎಂಬ ಸಣ್ಣ ಕೋಶವು ಅಂತಿಮವಾಗಿ ಪತ್ರರಂಧ್ರವನ್ನು ಸುತ್ತುವರೆದಿರುವ ಕಾವಲು ಕೋಶಗಳಾಗಿ ಪ್ರತ್ಯೇಕಿಸುತ್ತದೆ. ಈ ಮೆರಿಸ್ಟೆಮೊಯ್ಡ್ ನಂತರ ಅಸಮಪಾರ್ಶ್ವವಾಗಿ ಒಂದರಿಂದ ಮೂರು ಬಾರಿ ವಿಭಜಿಸುವ ಮೊದಲು ಕಾವಲುಗಾರ ತಾಯಿಯ ಕೋಶವಾಗಿ ವಿಭಜಿಸುತ್ತದೆ. ಕಾವಲುಗಾರ ತಾಯಿ ಕೋಶವು ನಂತರ ಒಂದು ಸಮ್ಮಿತೀಯ ವಿಭಾಗವನ್ನು ಮಾಡುತ್ತದೆ, ಇದು ಒಂದು ಜೋಡಿ ಕಾವಲು ಕೋಶಗಳನ್ನು ರೂಪಿಸುತ್ತದೆ.[೧೭] ಕೋಶ ವಿಭಜನೆಯು ಕೆಲವು ಜೀವಕೋಶಗಳಲ್ಲಿ ಪ್ರತಿಬಂಧಿಸುತ್ತದೆ ಆದ್ದರಿಂದ ಪತ್ರರಂಧ್ರ ನಡುವೆ ಯಾವಾಗಲೂ ಕನಿಷ್ಠ ಒಂದು ಕೋಶ ಇರುತ್ತದೆ.[೧೮]
ಇಪಿಎಫ್ (ಎಪಿಡರ್ಮಲ್ ಪ್ಯಾಟರ್ನಿಂಗ್ ಫ್ಯಾಕ್ಟರ್), ಇಆರ್ಎಲ್ (ಇರೆಕ್ಟಾ ಲೈಕ್) ಮತ್ತು ಯೋಡಾ ( ಪ್ಯೂಟೇಟಿವ್ ಮ್ಯಾಪ್ ಕೈನೇಸ್ ಕೈನೇಸ್ ಕೈನೇಸ್ ) ನಂತಹ ಅನೇಕ ಸಿಗ್ನಲ್ ಟ್ರಾನ್ಸ್ಡಕ್ಷನ್ ಘಟಕಗಳ ಪರಸ್ಪರ ಕ್ರಿಯೆಯಿಂದ ಪತ್ರರಂಧ್ರದ ವಿನ್ಯಾಸವನ್ನು ನಿಯಂತ್ರಿಸಲಾಗುತ್ತದೆ.[೧೮] ಈ ಅಂಶಗಳನ್ನು ಎನ್ಕೋಡ್ ಮಾಡುವ ಯಾವುದೇ ಜೀನ್ಗಳಲ್ಲಿನ ರೂಪಾಂತರಗಳು ಎಪಿಡರ್ಮಿಸ್ನಲ್ಲಿ ಪತ್ರರಂಧ್ರದ ಬೆಳವಣಿಗೆಯನ್ನು ಬದಲಾಯಿಸಬಹುದು.[೧೮] ಉದಾಹರಣೆಗೆ, ಒಂದು ಜೀನ್ನಲ್ಲಿನ ರೂಪಾಂತರವು ಹೆಚ್ಚು ಪತ್ರರಂಧ್ರವನ್ನು ಒಟ್ಟಿಗೆ ಜೋಡಿಸಲು ಕಾರಣವಾಗುತ್ತದೆ, ಆದ್ದರಿಂದ ಇದನ್ನು ಹಲವಾರು ಬಾಯಿಗಳು ( TMM ) ಎಂದು ಕರೆಯಲಾಗುತ್ತದೆ.[೧೭] ಆದರೆ, ಎಸ್ಪಿಸಿಎಚ್ ಜೀನ್ನ ಅಡ್ಡಿಯು ಒಟ್ಟಾಗಿ ಪತ್ರರಂಧ್ರದ ಬೆಳವಣಿಗೆಯನ್ನು ತಡೆಯುತ್ತದೆ.[೧೮] ಪತ್ರರಂಧ್ರದ ಉತ್ಪಾದನೆಯ ಸಕ್ರಿಯಗೊಳಿಸುವಿಕೆಯು ಇಪಿಎಫ಼್೧ ನ ಸಕ್ರಿಯಗೊಳಿಸುವಿಕೆಯಿಂದ ಸಂಭವಿಸಬಹುದು, ಇದು ಟಿಎಮ್ಎಮ್ ಅನ್ನು ಸಕ್ರಿಯಗೊಳಿಸುತ್ತದೆ, ಇದು ಒಟ್ಟಿಗೆ ಯೊಡ ಅನ್ನು ಸಕ್ರಿಯಗೊಳಿಸುತ್ತದೆ. ಎಸ್ಪಿಸಿಎಚ್ ಚಟುವಟಿಕೆಯನ್ನು ಕಡಿಮೆ ಮಾಡುತ್ತದೆ, ಇದು ಪತ್ರರಂಧ್ರದ ರಚನೆಯನ್ನು ಪ್ರಾರಂಭಿಸುವ ಅಸಮಪಾರ್ಶ್ವದ ಕೋಶ ವಿಭಜನೆಗೆ ಅನುವು ಮಾಡಿಕೊಡುತ್ತದೆ.[೧೮][೧೯] ಪತ್ರರಂಧ್ರದ ಬೆಳವಣಿಗೆಯು ಸ್ಟೊಮಾಜೆನ್ ಎಂಬ ಸೆಲ್ಯುಲಾರ್ ಪೆಪ್ಟೈಡ್ ಸಿಗ್ನಲ್ನಿಂದ ಸಹ ಸಂಯೋಜಿಸಲ್ಪಟ್ಟಿದೆ, ಇದು SPCH ನ ಪ್ರತಿಬಂಧವನ್ನು ಸಂಕೇತಿಸುತ್ತದೆ, ಇದರಿಂದಾಗಿ ಪತ್ರರಂಧ್ರದ ಸಂಖ್ಯೆ ಹೆಚ್ಚಾಗುತ್ತದೆ.[೨೦]
ಪರಿಸರ ಮತ್ತು ಹಾರ್ಮೋನುಗಳ ಅಂಶಗಳು ಸ್ಟೊಮಾಟಲ್ ಬೆಳವಣಿಗೆಯ ಮೇಲೆ ಪರಿಣಾಮ ಬೀರಬಹುದು. ಬೆಳಕು ಸಸ್ಯಗಳಲ್ಲಿ ಪತ್ರರಂಧ್ರದ ಬೆಳವಣಿಗೆಯನ್ನು ಹೆಚ್ಚಿಸುತ್ತದೆ; ಅದೇ ಸಮಯದಲ್ಲಿ, ಕತ್ತಲೆಯಲ್ಲಿ ಬೆಳೆದ ಸಸ್ಯಗಳು ಕಡಿಮೆ ಪ್ರಮಾಣದ ಪತ್ರರಂಧ್ರವನ್ನು ಹೊಂದಿರುತ್ತವೆ. ಇಆರ್ಎಲ್ ಮತ್ತು ಟಿಎಮ್ಎಮ್ ಗ್ರಾಹಕಗಳಂತಹ ಗ್ರಾಹಕ ಮಟ್ಟದಲ್ಲಿ ಅವುಗಳ ಬೆಳವಣಿಗೆಯ ಮೇಲೆ ಪರಿಣಾಮ ಬೀರುವ ಮೂಲಕ ಆಕ್ಸಿನ್ ಸ್ಟೊಮಾಟಲ್ ಬೆಳವಣಿಗೆಯನ್ನು ನಿಗ್ರಹಿಸುತ್ತದೆ. ಆದಾಗ್ಯೂ, ಆಕ್ಸಿನ್ನ ಕಡಿಮೆ ಸಾಂದ್ರತೆಯು ಕಾವಲುಗಾರ ತಾಯಿಯ ಜೀವಕೋಶದ ಸಮಾನ ವಿಭಜನೆಗೆ ಅನುವು ಮಾಡಿಕೊಡುತ್ತದೆ ಮತ್ತು ಕಾವಲು ಕೋಶಗಳನ್ನು ಉತ್ಪಾದಿಸುವ ಅವಕಾಶವನ್ನು ಹೆಚ್ಚಿಸುತ್ತದೆ.[೨೧]
ಹೆಚ್ಚಿನ ಆಂಜಿಯೋಸ್ಪರ್ಮ್ ಮರಗಳು ತಮ್ಮ ಕೆಳಗಿನ ಎಲೆಯ ಮೇಲ್ಮೈಯಲ್ಲಿ ಮಾತ್ರ ಸ್ಟೊಮಾಟಾವನ್ನು ಹೊಂದಿರುತ್ತವೆ. ಪಾಪ್ಲರ್ಗಳು ಮತ್ತು ವಿಲೋಗಳು ಅವುಗಳನ್ನು ಎರಡೂ ಮೇಲ್ಮೈಗಳಲ್ಲಿ ಹೊಂದಿರುತ್ತವೆ. ಎಲೆಗಳು ಎರಡೂ ಎಲೆಗಳ ಮೇಲ್ಮೈಗಳಲ್ಲಿ ಪತ್ರರಂಧ್ರವನ್ನು ಅಭಿವೃದ್ಧಿಪಡಿಸಿದಾಗ, ಕೆಳಗಿನ ಮೇಲ್ಮೈಯಲ್ಲಿರುವ ಪತ್ರರಂದ್ರವು ದೊಡ್ಡದಾಗಿರುತ್ತವೆ ಮತ್ತು ಹೆಚ್ಚು ಸಂಖ್ಯೆಯಲ್ಲಿರುತ್ತವೆ, ಆದರೆ ಜಾತಿಗಳು ಮತ್ತು ಜೀನೋಟೈಪ್ಗಳ ಬಗ್ಗೆ ಗಾತ್ರ ಮತ್ತು ಆವರ್ತನದಲ್ಲಿ ದೊಡ್ಡ ಮಟ್ಟದ ವ್ಯತ್ಯಾಸವಿರಬಹುದು. ಬಿಳಿ ಬೂದಿ ಮತ್ತು ಬಿಳಿ ಬರ್ಚ್ ಎಲೆಗಳು ಕಡಿಮೆ ಸ್ಟೊಮಾಟಾವನ್ನು ಹೊಂದಿದ್ದರು ಅವುಗಳ ಗಾತ್ರ ದೊಡ್ಡದಾಗಿದೆ. ಮತ್ತೊಂದೆಡೆ, ಸಕ್ಕರೆ ಮೇಪಲ್ ಮತ್ತು ಸಿಲ್ವರ್ ಮೇಪಲ್ ಸಣ್ಣ ಸ್ಟೊಮಾಟಾವನ್ನು ಹೊಂದಿದ್ದರು, ಅವುಗಳು ಹೆಚ್ಚು ಸಂಖ್ಯೆಯಲ್ಲಿವೆ.[೨೨]
ವಿಧಗಳು
ಪತ್ರರಂಧ್ರ ವಿಧಗಳ ವಿವಿಧ ವರ್ಗೀಕರಣಗಳು ಅಸ್ತಿತ್ವದಲ್ಲಿವೆ. ೧೮೮೬ ರಲ್ಲಿ ಜೂಲಿಯನ್ ಜೋಸೆಫ್ ವೆಸ್ಕ್ ಪರಿಚಯಿಸಿದ ಪ್ರಕಾರಗಳನ್ನು ಆಧರಿಸಿ ವ್ಯಾಪಕವಾಗಿ ಬಳಸಲಾಗುತ್ತದೆ, ಇದನ್ನು ಮೆಟ್ಕಾಲ್ಫ್ ಮತ್ತು ಚಾಕ್ನಿಂದ ಅಭಿವೃದ್ಧಿಪಡಿಸಲಾಯಿತು,[೨೩] ಮತ್ತು ನಂತರ ಇತರ ಲೇಖಕರು ಇದನ್ನು ಪೂರಕಗೊಳಿಸಿದರು. ಇದು ಎರಡು ಕಾವಲು ಕೋಶಗಳನ್ನು ಸುತ್ತುವರೆದಿರುವ ಅಂಗ ಕೋಶಗಳ ಗಾತ್ರ, ಆಕಾರ ಮತ್ತು ಜೋಡಣೆಯನ್ನು ಆಧರಿಸಿದೆ.[೨೪] ಅವರು ಡಿಕೋಟ್ಗಳಿಗೆ ಪ್ರತ್ಯೇಕಿಸುತ್ತಾರೆ:
- ಆಕ್ಟಿನೊಸೈಟಿಕ್ಆಕ್ಟಿನೊಸೈಟಿಕ್ ( ನಕ್ಷತ್ರ-ಕೋಶ) ಪತ್ರರಂಧ್ರವು ಕಾವಲು ಕೋಶಗಳನ್ನು ಹೊಂದಿದ್ದು ಅದು ನಕ್ಷತ್ರದಂತಹ ವೃತ್ತವನ್ನು ರೂಪಿಸುವ ಕನಿಷ್ಠ ಐದು ವಿಕಿರಣ ಕೋಶಗಳಿಂದ ಆವೃತವಾಗಿದೆ. ಇದು ಅಪರೂಪದ ವಿಧವಾಗಿದ್ದು, ಉದಾಹರಣೆಗೆ ಎಬೆನೇಸಿಯ ಕುಟುಂಬದಲ್ಲಿ ಕಂಡುಬರುತ್ತದೆ.
- ಅನಿಸೊಸೈಟಿಕ್ಅನಿಸೊಸೈಟಿಕ್ ( ಅಸಮಾನ ಕೋಶ) ಪತ್ರರಂಧ್ರವು ಎರಡು ದೊಡ್ಡ ಅಂಗ ಕೋಶಗಳ ಮತ್ತು ಒಂದು ಸ್ಪಷ್ಟವಾಗಿ ಚಿಕ್ಕದಾದ ನಡುವೆ ಕಾವಲು ಕೋಶಗಳನ್ನು ಹೊಂದಿರುತ್ತದೆ. ಈ ರೀತಿಯ ಸ್ಟೊಮಾಟಾವನ್ನು ಬ್ರಾಸಿಕೇಸಿ, ಸೋಲನೇಸಿ ಮತ್ತು ಕ್ರಾಸ್ಸುಲೇಸಿ ಸೇರಿದಂತೆ ಮೂವತ್ತಕ್ಕೂ ಹೆಚ್ಚು ಡಿಕಾಟ್ ಕುಟುಂಬಗಳಲ್ಲಿ ಕಾಣಬಹುದು. ಇದನ್ನು ಕೆಲವೊಮ್ಮೆ ಕ್ರೂಸಿಫೆರಸ್ ಪ್ರಕಾರ ಎಂದು ಕರೆಯಲಾಗುತ್ತದೆ.
- ಅನೋಮೊಸೈಟಿಕ್ ಅನೋಮೊಸೈಟಿಕ್( ಅನಿಯಮಿತ ಕೋಶ) ಪತ್ರರಂಧ್ರವು ಕೋಶಗಳಿಂದ ಸುತ್ತುವರೆದಿರುವ ಕಾವಲು ಕೋಶಗಳನ್ನು ಹೊಂದಿರುತ್ತದೆ, ಅದು ಉಳಿದ ಎಪಿಡರ್ಮಿಸ್ ಕೋಶಗಳಂತೆಯೇ ಅದೇ ಗಾತ್ರ, ಆಕಾರ ಮತ್ತು ಜೋಡಣೆಯನ್ನು ಹೊಂದಿರುತ್ತದೆ. ಈ ರೀತಿಯ ಪತ್ರರಂಧ್ರವನ್ನು ನೂರಕ್ಕೂ ಹೆಚ್ಚು ಡಿಕೋಟ್ ಕುಟುಂಬಗಳಾದ ಅಪೊಸಿನೇಸಿ, ಬೋರಜಿನೇಸಿ, ಚೆನೊಪೊಡಿಯಾಸಿ ಮತ್ತು ಕ್ಯುಕುರ್ಬಿಟೇಸಿಯಂತಹ ಕುಟುಂಬಗಳಲ್ಲಿ ಕಾಣಬಹುದು. ಇದನ್ನು ಕೆಲವೊಮ್ಮೆ ರಾನ್ಕ್ಯುಲೇಶಿಯಸ್ ವಿಧ ಎಂದು ಕರೆಯಲಾಗುತ್ತದೆ.
- ಡಯಾಸಿಟಿಕ್ ಡಯಾಸಿಟಿಕ್(ಅಂದರೆ ಅಡ್ಡ-ಕೋಶೀಯ ) ಪತ್ರರಂಧ್ರವು ಎರಡು ಅಂಗ ಕೋಶಗಳಿಂದ ಸುತ್ತುವರೆದಿರುವ ಕಾವಲು ಕೋಶಗಳನ್ನು ಹೊಂದಿರುತ್ತದೆ, ಪ್ರತಿಯೊಂದೂ ತೆರೆಯುವಿಕೆಯ ಒಂದು ತುದಿಯನ್ನು ಸುತ್ತುವರಿಯುತ್ತದೆ ಮತ್ತು ತೆರೆಯುವಿಕೆಯ ಮಧ್ಯದ ಎದುರು ಪರಸ್ಪರ ಸಂಪರ್ಕಿಸುತ್ತದೆ. ಕ್ಯಾರಿಯೋಫಿಲೇಸಿ ಮತ್ತು ಅಕಾಂಥೇಸಿಯಂತಹ ಹತ್ತಕ್ಕೂ ಹೆಚ್ಚು ಡಿಕೋಟ್ ಕುಟುಂಬಗಳಲ್ಲಿ ಈ ರೀತಿಯ ಪತ್ರರಂಧ್ರವನ್ನು ಕಾಣಬಹುದು. ಇದನ್ನು ಕೆಲವೊಮ್ಮೆ ಕ್ಯಾರಿಯೋಫಿಲೇಸಿಯಸ್ ಪ್ರಕಾರ ಎಂದು ಕರೆಯಲಾಗುತ್ತದೆ.
- ಹೆಮಿಪ್ಯಾರಾಸಿಟಿಕ್ಹೆಮಿಪ್ಯಾರಾಸಿಟಿಕ್ ಪತ್ರರಂಧ್ರವು ಕೇವಲ ಒಂದು ಸಹಾಯಕ ಕೋಶದಿಂದ ಗಡಿಯಾಗಿದೆ, ಇದು ಸುತ್ತಮುತ್ತಲಿನ ಎಪಿಡರ್ಮಿಸ್ ಕೋಶಗಳಿಂದ ಭಿನ್ನವಾಗಿರುತ್ತದೆ. ಅದರ ಉದ್ದವು ಸ್ಟೊಮಾ ತೆರೆಯುವಿಕೆಗೆ ಸಮಾನಾಂತರವಾಗಿರುತ್ತದೆ. ಈ ಪ್ರಕಾರವು ಮೊಲ್ಲುಜಿನೇಸಿ ಮತ್ತು ಐಜೋಯೇಸಿಯಲ್ಲಿ ಕಂಡುಬರುತ್ತದೆ.
- ಪ್ಯಾರಾಸೈಟಿಕ್ಪ್ಯಾರಾಸೈಟಿಕ್ ( ಸಮಾನಾಂತರ ಕೋಶ) ಪತ್ರರಂಧ್ರವು ಒಂದು ಅಥವಾ ಹೆಚ್ಚಿನ ಅಂಗ ಕೋಶಗಳನ್ನು ಕಾವಲು ಕೋಶಗಳ ನಡುವಿನ ತೆರೆಯುವಿಕೆಗೆ ಸಮಾನಾಂತರವಾಗಿ ಹೊಂದಿರುತ್ತದೆ. ಈ ಅಂಗ ಕೋಶಗಳು ಕಾವಲು ಕೋಶಗಳನ್ನು ಮೀರಿ ತಲುಪಬಹುದು ಅಥವಾ ಇಲ್ಲ. ಈ ರೀತಿಯ ಪತ್ರರಂಧ್ರವನ್ನು ನೂರಕ್ಕೂ ಹೆಚ್ಚು ಡಿಕೋಟ್ ಕುಟುಂಬಗಳಾದ ರೂಬಿಯೇಸಿ, ಕನ್ವೋಲ್ವುಲೇಸಿ ಮತ್ತು ಫ್ಯಾಬೇಸಿಯಲ್ಲಿ ಕಾಣಬಹುದು. ಇದನ್ನು ಕೆಲವೊಮ್ಮೆ ರೂಬಿಯಾಸಿಯಸ್ ವಿಧ ಎಂದು ಕರೆಯಲಾಗುತ್ತದೆ.
ಮೊನೊಕಾಟ್ಗಳಲ್ಲಿ, ಹಲವಾರು ರೀತಿಯ ಪತ್ರರಂಧ್ರಗಳು ಸಂಭವಿಸುತ್ತವೆ:
- ಗ್ರ್ಯಾಮಿನಸ್ ಅಥವಾ ಗ್ರ್ಯಾಮಿನಾಯ್ಡ್ ( ಹುಲ್ಲಿನಂತಿರುವ ಅರ್ಥ) ಪತ್ರರಂಧ್ರವು ಎರಡು ಮಸೂರ-ಆಕಾರದ ಅಂಗ ಕೋಶಗಳಿಂದ ಸುತ್ತುವರೆದಿರುವ ಎರಡು ಕಾವಲು ಕೋಶಗಳನ್ನು ಹೊಂದಿರುತ್ತದೆ. ಕಾವಲು ಕೋಶಗಳು ಮಧ್ಯದಲ್ಲಿ ಕಿರಿದಾಗಿರುತ್ತವೆ ಮತ್ತು ಪ್ರತಿ ತುದಿಯಲ್ಲಿ ಉಬ್ಬಿರುತ್ತವೆ. ಈ ಮಧ್ಯದ ವಿಭಾಗವು ಬಲವಾಗಿ ದಪ್ಪವಾಗಿರುತ್ತದೆ. ಅಂಗ ಕೋಶಗಳ ಅಕ್ಷವು ಸಮಾನಾಂತರ ಪತ್ರರಂಧ್ರ ತೆರೆಯುವಿಕೆಯಾಗಿದೆ. ಪೋಯೇಸಿ ಮತ್ತು ಸೈಪರೇಸಿ ಸೇರಿದಂತೆ ಮೊನೊಕಾಟ್ ಕುಟುಂಬಗಳಲ್ಲಿ ಈ ಪ್ರಕಾರವನ್ನು ಕಾಣಬಹುದು.[೨೫]
- ಹೆಕ್ಸಾಸಿಟಿಕ್ಹೆಕ್ಸಾಸಿಟಿಕ್ (ಅಂದರೆ ಆರು-ಕೋಶ ) ಪತ್ರರಂಧ್ರವು ಎರಡೂ ಕಾವಲು ಕೋಶಗಳ ಸುತ್ತಲೂ ಆರು ಸಹಾಯಕ ಕೋಶಗಳನ್ನು ಹೊಂದಿರುತ್ತದೆ, ಒಂದು ಪತ್ರರಂಧ್ರದ ತೆರೆಯುವಿಕೆಯ ಎರಡೂ ತುದಿಯಲ್ಲಿ, ಒಂದು ಪ್ರತಿ ಕಾವಲು ಕೋಶಕ್ಕೆ ಪಕ್ಕದಲ್ಲಿದೆ ಮತ್ತು ಆ ಕೊನೆಯ ಅಂಗ ಕೋಶ ಮತ್ತು ಪ್ರಮಾಣಿತ ಎಪಿಡರ್ಮಿಸ್ ಕೋಶಗಳ ನಡುವೆ ಒಂದು. ಈ ಪ್ರಕಾರವನ್ನು ಕೆಲವು ಮೊನೊಕೋಟ್ ಕುಟುಂಬಗಳಲ್ಲಿ ಕಾಣಬಹುದು.
- ಟೆಟ್ರಾಸೈಟಿಕ್ಟೆಟ್ರಾಸೈಟಿಕ್ (ಅಂದರೆ ನಾಲ್ಕು ಕೋಶಗಳ )ಪತ್ರರಂಧ್ರವು ನಾಲ್ಕು ಸಹಾಯಕ ಕೋಶಗಳನ್ನು ಹೊಂದಿರುತ್ತದೆ, ಒಂದು ತೆರೆಯುವಿಕೆಯ ಎರಡೂ ತುದಿಯಲ್ಲಿ ಮತ್ತು ಪ್ರತಿ ಕಾವಲು ಕೋಶದ ಪಕ್ಕದಲ್ಲಿ ಒಂದು. ಈ ವಿಧವು ಅನೇಕ ಮೊನೊಕಾಟ್ ಕುಟುಂಬಗಳಲ್ಲಿ ಕಂಡುಬರುತ್ತದೆ, ಆದರೆ ಟಿಲಿಯಾ ಮತ್ತು ಹಲವಾರು ಅಸ್ಕ್ಲೆಪಿಯಾಡೇಸಿಯಂತಹ ಕೆಲವು ಡಿಕಾಟ್ಗಳಲ್ಲಿಯೂ ಕಂಡುಬರುತ್ತದೆ.
ಜರೀಗಿಡಗಳಲ್ಲಿ, ನಾಲ್ಕು ವಿಭಿನ್ನ ಪ್ರಕಾರಗಳನ್ನು ಪ್ರತ್ಯೇಕಿಸಲಾಗಿದೆ:
- ಹೈಪೋಸೈಟಿಕ್ಹೈಪೋಸೈಟಿಕ್ ಸ್ಟೊಮಾಟಾವು ಸಾಮಾನ್ಯ ಎಪಿಡರ್ಮಿಸ್ ಕೋಶಗಳೊಂದಿಗೆ ಒಂದು ಪದರದಲ್ಲಿ ಎರಡು ಕಾವಲು ಕೋಶಗಳನ್ನು ಹೊಂದಿರುತ್ತದೆ, ಆದರೆ ಎಪಿಡರ್ಮಿಸ್ನ ಹೊರ ಮೇಲ್ಮೈಯಲ್ಲಿ ಎರಡು ಅಂಗ ಕೋಶಗಳನ್ನು ಹೊಂದಿರುತ್ತದೆ, ಕಾವಲು ಕೋಶಗಳಿಗೆ ಸಮಾನಾಂತರವಾಗಿ ಜೋಡಿಸಲಾಗಿದೆ, ಅವುಗಳ ನಡುವೆ ರಂಧ್ರವಿದೆ, ಸ್ಟೊಮಾ ತೆರೆಯುವಿಕೆಯ ಮೇಲೆ ಇರುತ್ತದೆ.
- ಪೆರಿಸೈಟಿಕ್ ಪೆರಿಸೈಟಿಕ್ ಪತ್ರರಂಧ್ರ ಎರಡು ಕಾವಲು ಕೋಶಗಳನ್ನು ಹೊಂದಿದ್ದು ಅದು ಸಂಪೂರ್ಣವಾಗಿ ಒಂದು ನಿರಂತರ ಅಂಗ ಕೋಶದಿಂದ ಸುತ್ತುವರಿಯಲ್ಪಟ್ಟಿದೆ (ಡೋನಟ್ನಂತೆ).
- ಡೆಸ್ಮೋಸೈಟಿಕ್ಡೆಸ್ಮೋಸೈಟಿಕ್ ಪತ್ರರಂಧ್ರ ಎರಡು ಕಾವಲು ಕೋಶಗಳನ್ನು ಹೊಂದಿದ್ದು, ಅದರ ತುದಿಗಳನ್ನು ವಿಲೀನಗೊಳಿಸದ ಒಂದು ಅಂಗ ಕೋಶದಿಂದ ಸಂಪೂರ್ಣವಾಗಿ ಸುತ್ತುವರಿಯಲ್ಪಟ್ಟಿದೆ (ಸಾಸೇಜ್ನಂತೆ).
- ಪೊಲೊಸೈಟಿಕ್ಪೊಲೊಸೈಟಿಕ್ ಪತ್ರರಂಧ್ರ ಎರಡು ಕಾವಲು ಕೋಶಗಳನ್ನು ಹೊಂದಿದ್ದು, ಅವುಗಳು ಒಂದು ಅಂಗ ಕೋಶದಿಂದ ಹೆಚ್ಚಾಗಿ ಸುತ್ತುವರಿಯಲ್ಪಟ್ಟಿವೆ, ಆದರೆ ಸಾಮಾನ್ಯ ಎಪಿಡರ್ಮಿಸ್ ಕೋಶಗಳನ್ನು ಸಹ ಸಂಪರ್ಕಿಸುತ್ತವೆ (ಯು ಅಥವಾ ಹಾರ್ಸ್ಶೂ ನಂತಹ).
ರೋಗಕಾರಕ ಮಾರ್ಗಗಳಾಗಿ ಪತ್ರರಂಧ್ರ
ಪತ್ರರಂಧ್ರವು ಎಲೆಯಲ್ಲಿನ ಸ್ಪಷ್ಟ ರಂಧ್ರಗಳಾಗಿದ್ದು, ಸ್ವಲ್ಪ ಸಮಯದವರೆಗೆ ಊಹಿಸಿದಂತೆ, ರೋಗಕಾರಕಗಳು ಸವಾಲು ಮಾಡದೆ ಪ್ರವೇಶಿಸಬಹುದು. ಆದಾಗ್ಯೂ, ಪತ್ರರಂಧ್ರ ವಾಸ್ತವವಾಗಿ ಕೆಲವು ರೋಗಕಾರಕಗಳ ಉಪಸ್ಥಿತಿಯನ್ನು ಗ್ರಹಿಸುತ್ತದೆ ಎಂದು ಇತ್ತೀಚೆಗೆ ತೋರಿಸಲಾಗಿದೆ. ಆದಾಗ್ಯೂ, ಪ್ರಯೋಗದಲ್ಲಿ ಅರಾಬಿಡೋಪ್ಸಿಸ್ ಸಸ್ಯದ ಎಲೆಗಳಿಗೆ ವೈರಲ್ ಬ್ಯಾಕ್ಟೀರಿಯಾವನ್ನು ಅನ್ವಯಿಸುವುದರೊಂದಿಗೆ, ಬ್ಯಾಕ್ಟೀರಿಯಾವು ರಾಸಾಯನಿಕ ಕೊರೊನಾಟೈನ್ ಅನ್ನು ಬಿಡುಗಡೆ ಮಾಡಿತು, ಇದು ಕೆಲವು ಗಂಟೆಗಳಲ್ಲಿ ಪತ್ರರಂಧ್ರವನ್ನು ಮತ್ತೆ ತೆರೆಯುವಂತೆ ಮಾಡಿತು.[೨೬]
ಪತ್ರರಂಧ್ರ ಮತ್ತು ಹವಾಮಾನ ಬದಲಾವಣೆ
ಪರಿಸರ ಅಂಶಗಳಿಗೆ ಸ್ಟೊಮಾಟಾದ ಪ್ರತಿಕ್ರಿಯೆ
ಬರವು ಪತ್ರರಂಧ್ರ ತೆರೆಯುವಿಕೆಯನ್ನು ತಡೆಯುತ್ತದೆ, ಆದರೆ ಮಧ್ಯಮ ಬರವು ಸೋಯಾ ಬೀನ್ಸ್ನ ಪತ್ರರಂಧ್ರಮುಚ್ಚುವಿಕೆಯ ಮೇಲೆ ಗಮನಾರ್ಹ ಪರಿಣಾಮವನ್ನು ಬೀರಲಿಲ್ಲ. ಸ್ಟೊಮಾಟಲ್ ಮುಚ್ಚುವಿಕೆಯ ವಿಭಿನ್ನ ಕಾರ್ಯವಿಧಾನಗಳಿವೆ. ಕಡಿಮೆ ಆರ್ದ್ರತೆಯು ಟರ್ಗರ್ ನಷ್ಟವನ್ನು ಉಂಟುಮಾಡುವ ಕಾವಲು ಕೋಶಗಳನ್ನು ಒತ್ತಿಹೇಳುತ್ತದೆ, ಇದನ್ನು ಹೈಡ್ರೊಪಾಸಿವ್ ಮುಚ್ಚುವಿಕೆ ಎಂದು ಕರೆಯಲಾಗುತ್ತದೆ. ಹೈಡ್ರೋಆಕ್ಟಿವ್ ಮುಚ್ಚುವಿಕೆಯು ಬರಗಾಲದ ಒತ್ತಡದಿಂದ ಪರಿಣಾಮ ಬೀರುವ ಸಂಪೂರ್ಣ ಎಲೆಯಂತೆ ವ್ಯತಿರಿಕ್ತವಾಗಿದೆ, ಇದು ಹೆಚ್ಚಾಗಿ ಅಬ್ಸಿಸಿಕ್ ಆಮ್ಲದಿಂದ ಪ್ರಚೋದಿಸಲ್ಪಟ್ಟಿದೆ ಎಂದು ನಂಬಲಾಗಿದೆ.[೨೭]
ದ್ಯುತಿಸಂಶ್ಲೇಷಣೆ, ಸಸ್ಯ ಜಲ ಸಾರಿಗೆ ( ಕ್ಸೈಲೆಮ್ ) ಮತ್ತು ಅನಿಲ ವಿನಿಮಯವು ಸಸ್ಯಗಳ ಕಾರ್ಯನಿರ್ವಹಣೆಯಲ್ಲಿ ಮುಖ್ಯವಾದ ಪತ್ರರಂಧ್ರ ಕ್ರಿಯೆಯಿಂದ ನಿಯಂತ್ರಿಸಲ್ಪಡುತ್ತದೆ.[೨೮]
ಪತ್ರರಂಧ್ರವು ಬೆಳಕಿಗೆ ಸ್ಪಂದಿಸುತ್ತದೆ ಮತ್ತು ನೀಲಿ ಬೆಳಕು ಪತ್ರರಂಧ್ರ ಪ್ರತಿಕ್ರಿಯೆಯನ್ನು ಉಂಟುಮಾಡುವಲ್ಲಿ ಕೆಂಪು ಬೆಳಕಿನಂತೆ ಸುಮಾರು ೧೦ ಪಟ್ಟು ಪರಿಣಾಮಕಾರಿಯಾಗಿದೆ. ಬ್ಲೂ ಲೈಟ್ಗೆ ಪತ್ರರಂಧ್ರದ ಬೆಳಕಿನ ಪ್ರತಿಕ್ರಿಯೆಯು ಕ್ಲೋರೊಫಿಲ್ನಂತಹ ಇತರ ಎಲೆ ಘಟಕಗಳಿಂದ ಸ್ವತಂತ್ರವಾಗಿದೆ ಎಂದು ಸಂಶೋಧನೆ ಸೂಚಿಸುತ್ತದೆ. ಪೊಟ್ಯಾಸಿಯಮ್ನ ಸಾಕಷ್ಟು ಲಭ್ಯತೆ ಇದ್ದಲ್ಲಿ ಗಾರ್ಡ್ ಸೆಲ್ ಪ್ರೊಟೊಪ್ಲಾಸ್ಟ್ಗಳು ನೀಲಿ ಬೆಳಕಿನ ಅಡಿಯಲ್ಲಿ ಉಬ್ಬುತ್ತವೆ.[೨೯] ದ್ಯುತಿಸಂಶ್ಲೇಷಣೆ ಪ್ರಕ್ರಿಯೆಯು ಪ್ರಾರಂಭವಾಗುವ ಮೊದಲು, ಪೊಟ್ಯಾಸಿಯಮ್ ಸಾಂದ್ರತೆಯನ್ನು ಹೆಚ್ಚಿಸುವುದರಿಂದ ಬೆಳಿಗ್ಗೆ ಪತ್ರರಂಧ್ರ ತೆರೆಯುವಿಕೆಯನ್ನು ಹೆಚ್ಚಿಸಬಹುದು ಎಂದು ಹಲವಾರು ಅಧ್ಯಯನಗಳು ಬೆಂಬಲವನ್ನು ಕಂಡುಕೊಂಡಿವೆ, ಆದರೆ ನಂತರದ ದಿನದಲ್ಲಿ ಸುಕ್ರೋಸ್ ಪತ್ರರಂಧ್ರ ತೆರೆಯುವಿಕೆಯನ್ನು ನಿಯಂತ್ರಿಸುವಲ್ಲಿ ದೊಡ್ಡ ಪಾತ್ರವನ್ನು ವಹಿಸುತ್ತದೆ.[೩೦]
ಪತ್ರರಂಧ್ರ ಸಾಂದ್ರತೆ ಮತ್ತು ದ್ಯುತಿರಂಧ್ರ (ಪತ್ರರಂಧ್ರದ ಉದ್ದ) ವಾತಾವರಣದ CO 2 ಸಾಂದ್ರತೆ, ಬೆಳಕಿನ ತೀವ್ರತೆ, ಗಾಳಿಯ ಉಷ್ಣತೆ ಮತ್ತು ದ್ಯುತಿ ಅವಧಿ (ಹಗಲಿನ ಅವಧಿ) ನಂತಹ ಹಲವಾರು ಪರಿಸರ ಅಂಶಗಳ ಅಡಿಯಲ್ಲಿ ಬದಲಾಗುತ್ತದೆ.[೩೧][೩೨]
ಪತ್ರರಂಧ್ರ ಸಾಂದ್ರತೆಯನ್ನು ಕಡಿಮೆ ಮಾಡುವುದು ವಾತಾವರಣದ CO 2 ([CO 2 ] atm ) ಸಾಂದ್ರತೆಯ ಹೆಚ್ಚಳಕ್ಕೆ ಸಸ್ಯಗಳು ಪ್ರತಿಕ್ರಿಯಿಸುವ ಒಂದು ಮಾರ್ಗವಾಗಿದೆ.[೩೩] [CO 2 ] atm ಪ್ರತಿಕ್ರಿಯೆಯಲ್ಲಿನ ಬದಲಾವಣೆಗಳು ಯಾಂತ್ರಿಕವಾಗಿ ಕಡಿಮೆ ಅರ್ಥವಾಗಿದ್ದರೂ, ಈ ಪತ್ರರಂಧ್ರ ಪ್ರತಿಕ್ರಿಯೆಯು ಪ್ರಸ್ಥಭೂಮಿಗೆ ಪ್ರಾರಂಭವಾಗಿದೆ, ಅಲ್ಲಿ ಇದು ಶೀಘ್ರದಲ್ಲೇ ಸಸ್ಯಗಳಲ್ಲಿನ ಟ್ರಾನ್ಸ್ಪಿರೇಷನ್ ಮತ್ತು ದ್ಯುತಿಸಂಶ್ಲೇಷಣೆ ಪ್ರಕ್ರಿಯೆಗಳ ಮೇಲೆ ಪರಿಣಾಮ ಬೀರುತ್ತದೆ ಎಂದು ನಿರೀಕ್ಷಿಸಲಾಗಿದೆ.[೨೮][೩೪]
ಹವಾಮಾನ ಬದಲಾವಣೆಯ ಸಮಯದಲ್ಲಿ ಭವಿಷ್ಯದ ರೂಪಾಂತರಗಳು
[CO 2 ] atm ೨೧೦೦ ರ ವೇಳೆಗೆ ೫೦೦-೧೦೦೦ ಪಿಪಿಎಮ್ ಅನ್ನು ತಲುಪುತ್ತದೆ ಎಂದು ನಿರೀಕ್ಷಿಸಲಾಗಿದೆ.[೨೮] ಕಳೆದ ೪೦೦೦೦೦ ವರ್ಷಗಳಲ್ಲಿ ೯೬% ೨೮೦ ಪಿಪಿಎಮ್ CO 2 ಮಟ್ಟಕ್ಕಿಂತ ಕಡಿಮೆ ಅನುಭವವನ್ನು ಹೊಂದಿದೆ. ಈ ಅಂಕಿ ಅಂಶದಿಂದ, ಇಂದಿನ ಸಸ್ಯಗಳ ಜೀನೋಟೈಪ್ಗಳು ಅವುಗಳ ಪೂರ್ವ-ಕೈಗಾರಿಕಾ ಸಂಬಂಧಿಯಿಂದ ಭಿನ್ನವಾಗಿರುವುದು ಹೆಚ್ಚು ಸಂಭವನೀಯವಾಗಿದೆ.[೨೮]
ಜೀನ್ HIC (ಹೆಚ್ಚಿನ ಇಂಗಾಲದ ಡೈಆಕ್ಸೈಡ್) ಸಸ್ಯಗಳಲ್ಲಿ ಪತ್ರರಂಧ್ರದ ಬೆಳವಣಿಗೆಗೆ ನಕಾರಾತ್ಮಕ ನಿಯಂತ್ರಕವನ್ನು ಸಂಕೇತಿಸುತ್ತದೆ.[೩೫] ಅರಬಿಡೋಪ್ಸಿಸ್ ಥಾಲಿಯಾನಾವನ್ನು ಬಳಸಿಕೊಂಡು HIC ಜೀನ್ನ ಸಂಶೋಧನೆಯು ಪ್ರಬಲವಾದ ಆಲೀಲ್ನಲ್ಲಿ ಪತ್ರರಂಧ್ರದ ಯಾವುದೇ ಹೆಚ್ಚಳವನ್ನು ಕಂಡುಕೊಂಡಿಲ್ಲ, ಆದರೆ 'ವೈಲ್ಡ್ ಟೈಪ್' ರಿಸೆಸಿವ್ ಆಲೀಲ್ನಲ್ಲಿ ವಾತಾವರಣದಲ್ಲಿ ಹೆಚ್ಚುತ್ತಿರುವ CO 2 ಮಟ್ಟಗಳಿಗೆ ಪ್ರತಿಕ್ರಿಯೆಯಾಗಿ ದೊಡ್ಡ ಹೆಚ್ಚಳವನ್ನು ತೋರಿಸಿದೆ.[೩೫] ಈ ಅಧ್ಯಯನಗಳು CO 2 ಮಟ್ಟವನ್ನು ಬದಲಿಸಲು ಸಸ್ಯಗಳ ಪ್ರತಿಕ್ರಿಯೆಯನ್ನು ಹೆಚ್ಚಾಗಿ ತಳಿಶಾಸ್ತ್ರದಿಂದ ನಿಯಂತ್ರಿಸುತ್ತದೆ ಎಂದು ಸೂಚಿಸುತ್ತದೆ.
ಕೃಷಿ ಪರಿಣಾಮಗಳು
ಫ್ರೀ-ಏರ್ ಕಾರ್ಬನ್ ಡೈಆಕ್ಸೈಡ್ ಎನ್ರಿಚ್ಮೆಂಟ್ (ಫೇಸ್) ಪ್ರಯೋಗಗಳ ಸಮಯದಲ್ಲಿ CO 2 ರಸಗೊಬ್ಬರದ ಪರಿಣಾಮವಾಗಿ ವಾತಾವರಣದಲ್ಲಿ ಹೆಚ್ಚಿದ CO 2 ಮಟ್ಟಗಳು ದ್ಯುತಿಸಂಶ್ಲೇಷಣೆಯನ್ನು ಹೆಚ್ಚಿಸುತ್ತದೆ, ಉತ್ಕರ್ಷಣೆಯನ್ನು ಕಡಿಮೆ ಮಾಡುತ್ತದೆ ಮತ್ತು ನೀರಿನ ಬಳಕೆಯ ದಕ್ಷತೆಯನ್ನು ಹೆಚ್ಚಿಸುತ್ತದೆ (ಡಬ್ಲೂಯುಇ) ಎಂದು ಅಂದಾಜಿಸಲಾಗಿದೆ.[೩೩] ೫೫೦ ಪಿಪಿಎಮ್ನ CO 2ನಲ್ಲಿ ಬೆಳೆ ಇಳುವರಿಯಲ್ಲಿ ೫-೫೦% ಹೆಚ್ಚಳವನ್ನು ಊಹಿಸುವ ಪ್ರಯೋಗಗಳಿಂದ ಸಿಮ್ಯುಲೇಶನ್ಗಳೊಂದಿಗೆ ಹೆಚ್ಚಿದ ಜೀವರಾಶಿ ಪರಿಣಾಮಗಳಲ್ಲಿ ಒಂದಾಗಿದೆ.[೩೬] ಎಲೆಗಳ ದ್ಯುತಿಸಂಶ್ಲೇಷಣೆಯ ದರಗಳು ಸಿ೩ ಸಸ್ಯಗಳಲ್ಲಿ ೩೦-೫೦% ರಷ್ಟು ಮತ್ತು ಸಿ೪ ನಲ್ಲಿ ೧೦-೨೫% ರಷ್ಟು ದ್ವಿಗುಣಗೊಂಡ CO 2 ಮಟ್ಟಗಳ ಅಡಿಯಲ್ಲಿ ಹೆಚ್ಚಾಗುತ್ತದೆ ಎಂದು ತೋರಿಸಲಾಗಿದೆ.[೩೬] ಪ್ರತಿಕ್ರಿಯೆ ಕಾರ್ಯವಿಧಾನದ ಅಸ್ತಿತ್ವವು [CO 2 ] atm ಗೆ ಪ್ರತಿಕ್ರಿಯೆಯಾಗಿ ಒಂದು ಫಿನೋಟೈಪಿಕ್ ಪ್ಲಾಸ್ಟಿಟಿಯನ್ನು ಉಂಟುಮಾಡುತ್ತದೆ, ಅದು ಸಸ್ಯದ ಉಸಿರಾಟ ಮತ್ತು ಕಾರ್ಯದ ವಿಕಾಸದಲ್ಲಿ ಹೊಂದಾಣಿಕೆಯ ಲಕ್ಷಣವಾಗಿರಬಹುದು.[೨೮][೩೨]
ರೂಪಾಂತರದ ಸಮಯದಲ್ಲಿ ಪತ್ರರಂಧ್ರ ಹೇಗೆ ಕಾರ್ಯನಿರ್ವಹಿಸುತ್ತದೆ ಎಂಬುದನ್ನು ಊಹಿಸುವುದು ನೈಸರ್ಗಿಕ ಮತ್ತು ಕೃಷಿ ವ್ಯವಸ್ಥೆಗಳಿಗೆ ಸಸ್ಯ ವ್ಯವಸ್ಥೆಗಳ ಉತ್ಪಾದಕತೆಯನ್ನು ಅರ್ಥಮಾಡಿಕೊಳ್ಳಲು ಉಪಯುಕ್ತವಾಗಿದೆ.[೩೧] ಸಸ್ಯ ತಳಿಗಾರರು ಮತ್ತು ರೈತರು ಆಹಾರ ಭದ್ರತೆಯ ಸವಾಲುಗಳ ಮುಖಾಂತರ ನೈಸರ್ಗಿಕವಾಗಿ ವಿಕಸನಗೊಳ್ಳುವ ಶಾಖ ಮತ್ತು ಬರ ನಿರೋಧಕ ಬೆಳೆ ಪ್ರಭೇದಗಳಂತಹ ಅತ್ಯುತ್ತಮ ಸೂಕ್ತವಾದ ಜಾತಿಗಳನ್ನು ಕಂಡುಹಿಡಿಯಲು ವಿಕಸನೀಯ ಮತ್ತು ಭಾಗವಹಿಸುವಿಕೆಯ ಸಸ್ಯ ತಳಿಗಳನ್ನು ಬಳಸಿಕೊಂಡು ಒಟ್ಟಾಗಿ ಕೆಲಸ ಮಾಡಲು ಪ್ರಾರಂಭಿಸಿದ್ದಾರೆ.[೩೩]
ಉಲ್ಲೇಖಗಳು
- ↑ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ ೩.೦ ೩.೧ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ Debbie Swarthout and C.Michael Hogan. 2010. Stomata. Encyclopedia of Earth. National Council for Science and the Environment, Washington DC
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite web
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ೧೨.೦ ೧೨.೧ ೧೨.೨ ಟೆಂಪ್ಲೇಟು:Cite web
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Citation
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ ೧೭.೦ ೧೭.೧ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ೧೮.೦ ೧೮.೧ ೧೮.೨ ೧೮.೩ ೧೮.೪ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ ೨೮.೦ ೨೮.೧ ೨೮.೨ ೨೮.೩ ೨೮.೪ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ ಟೆಂಪ್ಲೇಟು:Cite book
- ↑ ೩೧.೦ ೩೧.೧ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ೩೨.೦ ೩೨.೧ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ೩೩.೦ ೩೩.೧ ೩೩.೨ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ೩೫.೦ ೩೫.೧ ಟೆಂಪ್ಲೇಟು:Cite journal
- ↑ ೩೬.೦ ೩೬.೧ ಟೆಂಪ್ಲೇಟು:Cite journal